

成都生物所揭示蛇类起源及表型演化的遗传机制
作者:彭长军
时间:2023-06-20
6月19日,中国科学院成都生物研究所李家堂研究团队选取了全球极具代表性的蛇类物种,整合谱系基因组学、大规模比较转录组学分析方法及基因编辑等实验技术,综合探讨了蛇类起源及表型演化的遗传机制。相关论文以“Large-scale snake genome analyses provide insights into vertebrate development”为题,发表于国际期刊《细胞》(Cell)。
全球蛇类约4,000种,广泛分布于除南极洲外的各大洲陆地和海洋,在进化历史上处于脊椎动物演化的关键节点,是脊椎动物中重要的类群。蛇类具有四肢缺失、身体延长、左右肺不对称发育等特殊表型,揭示这些特殊表型的遗传机制对理解脊椎动物演化具有重要意义。
该研究表明蛇类起源于约1.18亿年前早白垩纪,与帝王蛇蜥和科摩多巨蜥组成的支系互为姐妹枝,这提示蛇类的祖先起源于蜥蜴类物种。蛇类最先分化出的类群为盲蛇类物种(Typhlopidae 和 Leptotyphlopidae),在白垩纪-早第三纪分界点(K-Pg, 约6500万年前)的生物大灭绝事件之后迎来了物种的爆发。
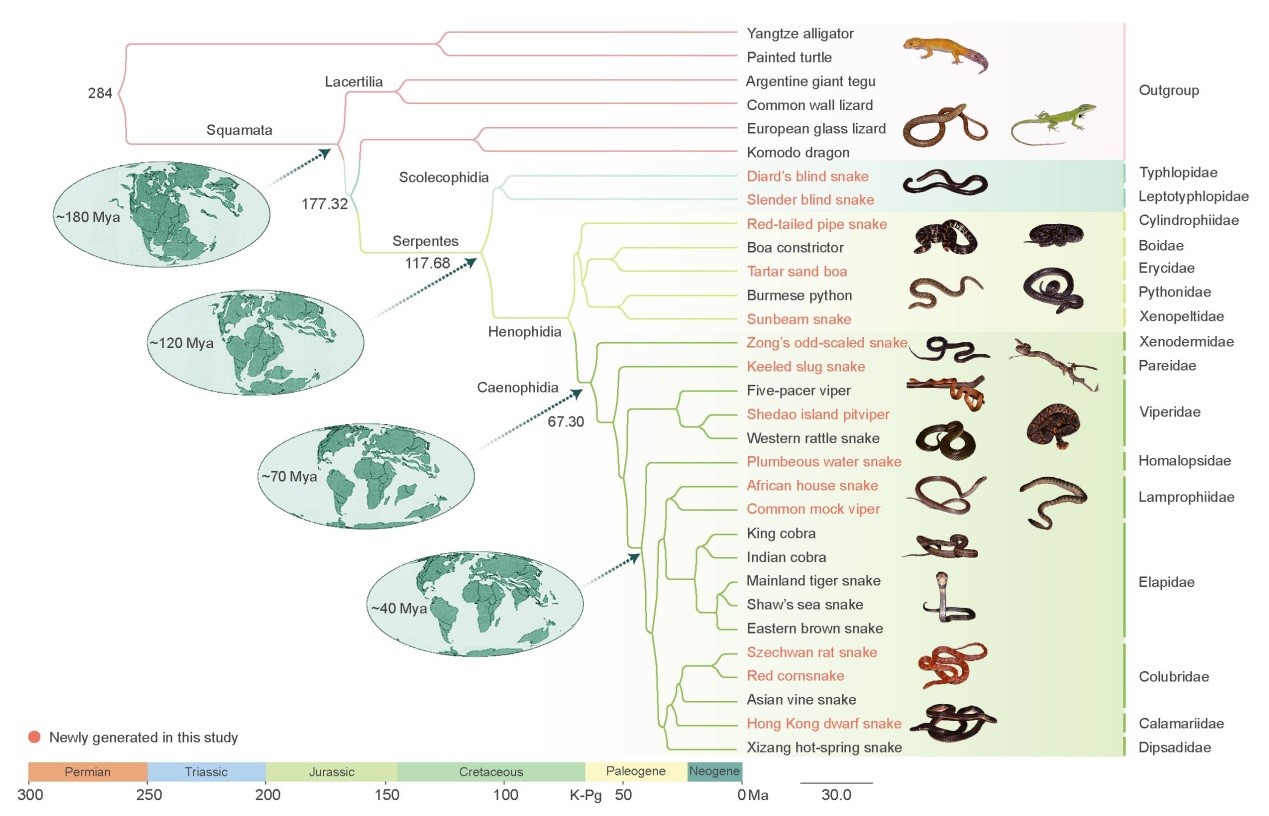
图1.全基因组序列极大似然法重构的31物种(6个外群和25种蛇)系统基因组学框架。
研究重构的蛇类祖先基因组由23对染色体组成,其中包含8对大染色体和15对小染色体。蛇类Z染色体可能起源于蜥蜴的常染色体且在整个蛇类演化中,至少发生过一次断裂。基因组大小与转座子含量的高低呈显著正相关关系,这与蛇类基因组大小与转座子含量无关的结论(之前基于短序列测序提示的结论)是不一致的。与鸟类、哺乳类动物相反,蛇类基因组的GC含量向着降低的方向演化,表明其基因组演化道路可能不同于其它脊椎动物类群。
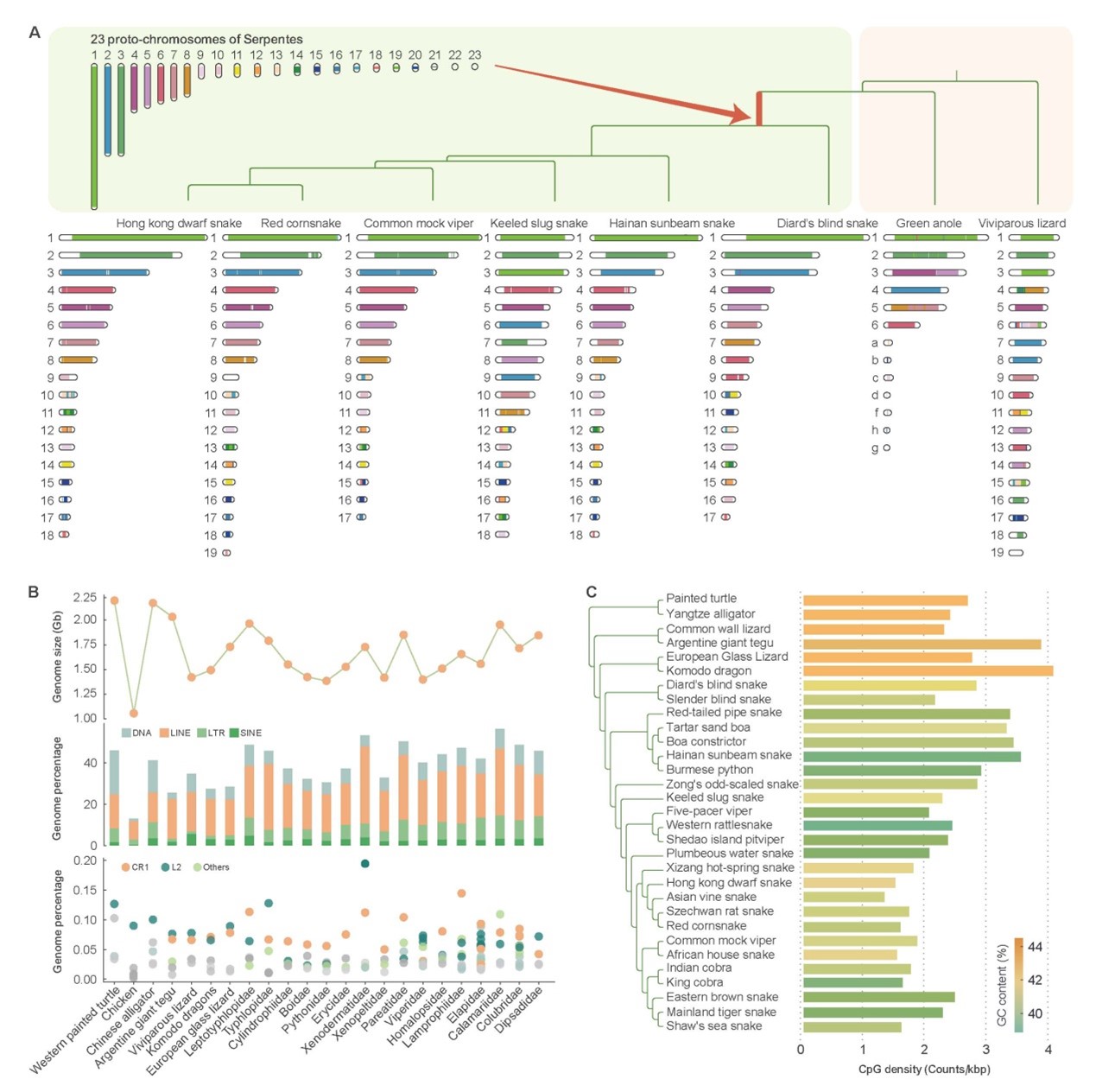
图2. 蛇类基因组演化特征。
蛇类可吞食超过自己头部大小的食物。该研究发现7个与头骨发育有关的正选择基因(HOX7, MMP14, ALX1, LIMK2, CHST11, KAT6B和TCOF1)可能是蛇类头骨特化可进行大幅吞食的重要遗传基础。与消化有关基因的丢失(GHRL, GHSR和MALR1)及适应性演化(GBA2和TRYP2),则可能使蛇类能够适应“暴饮暴食”的饮食习惯。
四肢缺失是蛇类最为特殊的特征。该研究揭示除ZRS增强子外,PTCH1基因编码蛋白的三个氨基酸残基缺失,可能是蛇类四肢缺失的重要遗传机制之一。随着四肢缺失,蛇类的体节数增加,身体延长。研究发现控制脊椎前端发育(FOXC2)和后端发育(DLLC)的重要基因受到强烈正选择,提示它们可能是蛇类身体延长的重要遗传基础。蛇类为适应身体延长,部分器官不对称发育,如退化了一侧肺。研究发现蛇类丢失了控制对称发育的DNAH11和FXJ1B基因,这可能是蛇类肺左右不对称发育的关键原因,研究成果对理解脊椎动物的器官不对称发育具有重要意义。
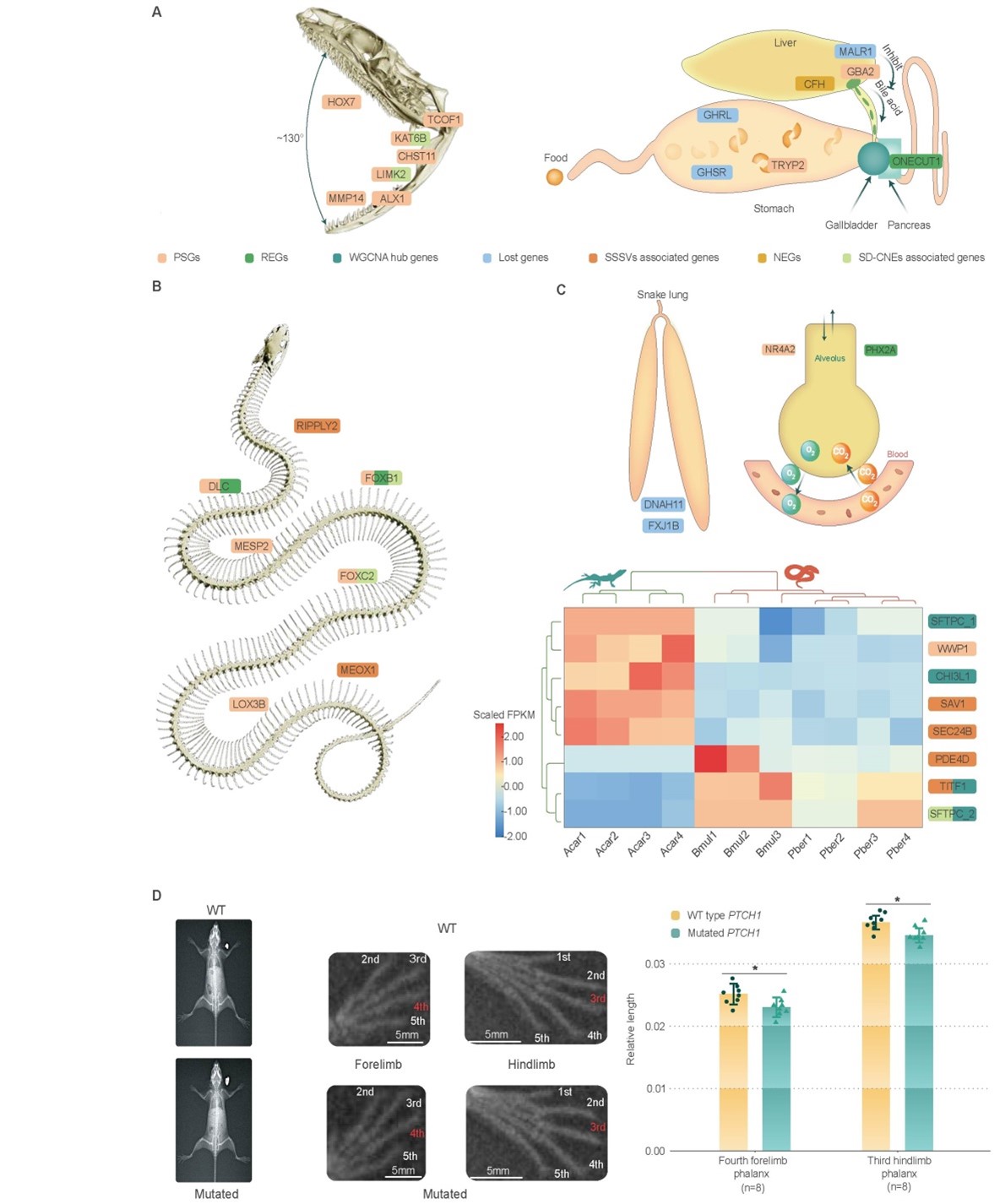
图3. 蛇类骨骼系统演化及器官适应性机制。
蛇类无活动眼睑,眼结构简化,视觉退化。研究表明蛇类丢失了8个光受体相关编码基因和3个对晶状体发育至关重要的基因(CRBB3, CRBB1和 CRBA1)。比较转录组分析进一步揭示,相比较于安乐蜥,蛇类15个光受体细胞维持相关基因显著上调表达。其中ESRRB的上调表达可能受到蛇类基因组特异结构变异的影响,6个基因(PDE6G, AGTPBP1, 2个CLTCs, MKKS和NXNL1)的上调表达可能与其调控元件的快速演化有关。这些提示蛇类可能经历了一个缺少光照的时期,而后又再次适应了明亮环境,调控元件的快速演化可能促进了这一适应过程。
蛇类虽然不是聋子,但没有外耳、中耳,其听力范围仅为50-1,000 Hz,远小于人类(10-20,000Hz)和蜥蜴(<100-7,700 Hz)。研究发现,16个蛇类耳发育相关基因的潜在调控元件发生了快速演化,重要耳蜗蛋白(otospiralin)编码基因在蛇类中丢失,表明它们可能是蛇类耳形态变化的遗传基础。此外,感受机械振动相关的基因(ASIC2)受到强烈正选择,且在蛇类内耳高表达,揭示它可能是蛇类对机械振动敏感的主要原因。尽管蛇类是否有味觉存在争议,该研究发现蛇类普遍拥有味觉受体及味觉传导相关信号通路,且苦味受体编码基因(TAS2R40)快速演化,提示蛇类的味觉可能并未丢失,且功能可能有一定强化。
盲蛇类物种是蛇类中高度特化的穴居类群,其头骨高度愈合,眼、牙高度退化,专食蚂蚁及蚂蚁卵。该研究发现与非盲蛇相比,47个编码基因在盲蛇中特异丢失,其中包括8个视觉相关基因。比较转录组分析发现12个视感受相关的基因在盲蛇眼中显著低表达,这些基因组元件的改变可能导致盲蛇类视觉丧失。此外,该研究发现,盲蛇与骨骼及牙齿发育相关编码基因的调控元件快速演化,可能是盲蛇头骨特化及牙齿退化的主要原因。盲蛇适应穴居生活,专食富含几丁质的蚂蚁及蚂蚁卵。研究发现,几丁质酶编码基因受到强烈正选择。酶活性检测实验表明,大盲蛇中该几丁质酶的催化活性显著高于缅甸蟒。这表明盲蛇可通过增强几丁质酶的催化活性,消化蚂蚁及蚂蚁卵中含量颇高的几丁质。
对红外光敏感的蟒蚺类蛇和蝮蛇以其特殊的红外感受器官唇窝和颊窝而闻名于世。研究发现,与热响应有关的PMP22基因及与三叉神经发育相关的NFIB基因的调控元件在这两类蛇中发生了趋同快速演化,提示非编码调控元件的快速演化是部分蛇类红外感受能力产生的遗传基础,这对综合理解脊椎动物红外感应的机制具有重要意义。
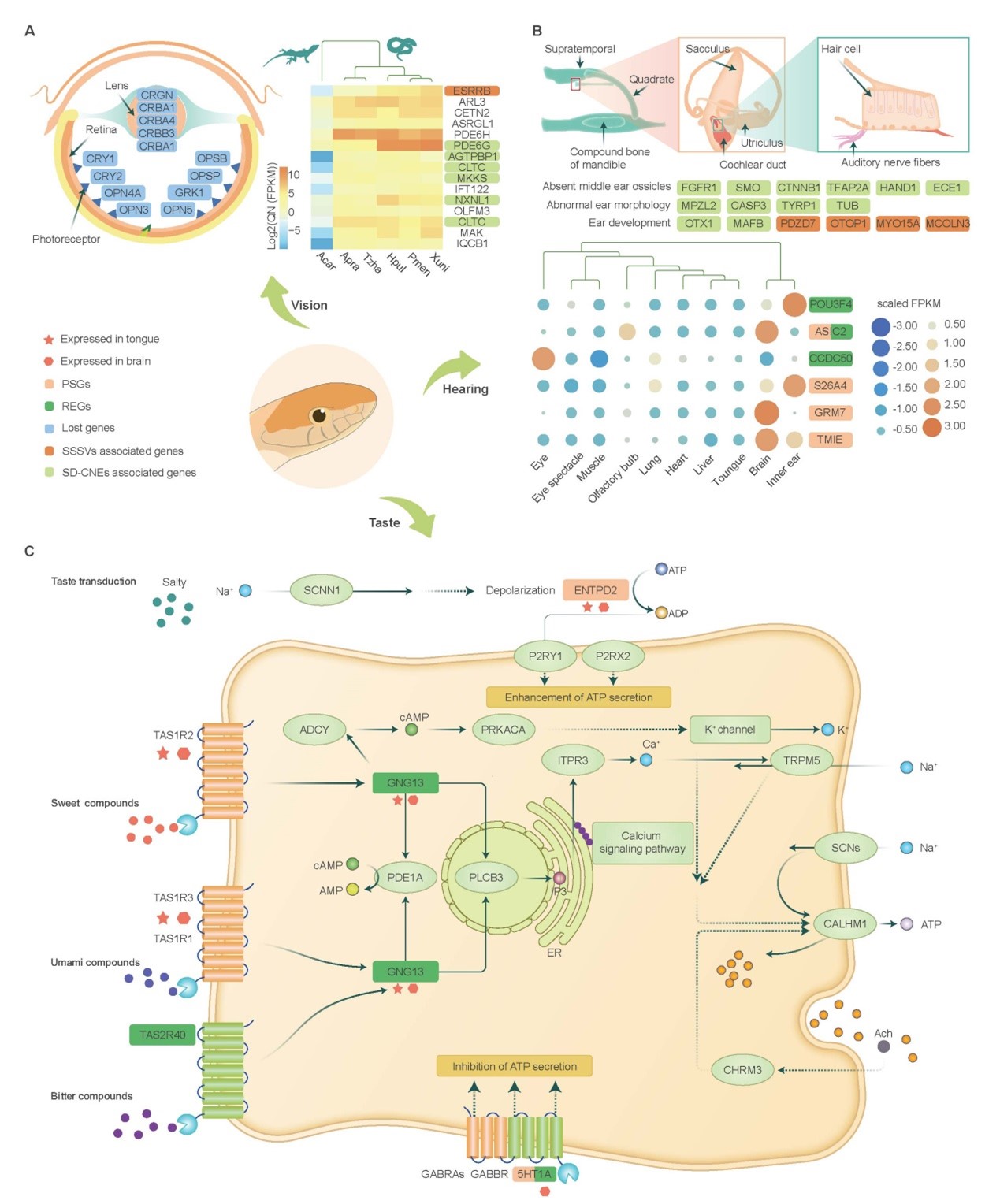
图4. 蛇类感官系统演化。
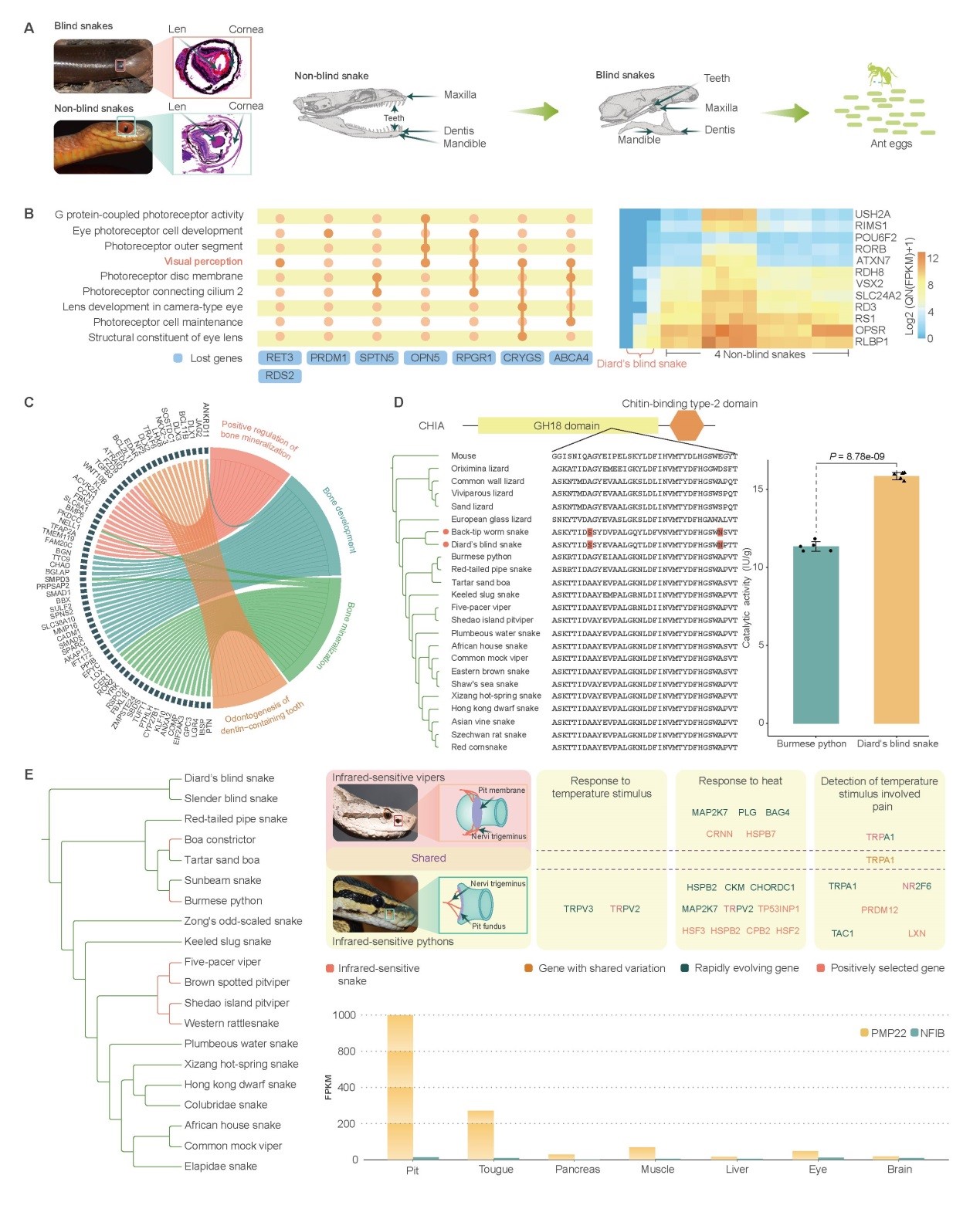
图5.盲蛇和红外感应蛇类的演化。
该研究率先启动了全球尺度下的蛇类大规模组学研究,构建了蛇类最有力的系统基因组学框架,阐明了蛇类的起源及其特殊表型如四肢缺失、肺不对称发育等表型特征背后的遗传机制。上述研究对理解脊椎动物演化历史具有重要意义,将推动动物演化生物学等相关学科的发展。
李家堂团队未来将聚焦开发玉米蛇为模式动物并开展演化发育生物学研究。同时,围绕蛇毒等重要遗传资源的挖掘和运用,为抗蛇毒血清及蛇毒衍生药物的研发提供科学支撑。研究团队将推动从基础科学到应用基础科学的发展,更好的服务国家重大战略需求。
中国科学院成都生物研究所博士生彭长军、昆明动物研究所吴东东研究员和成都生物研究所助理研究员任金龙为该论文共同第一作者,成都生物研究所李家堂研究员为该论文的独立通讯作者。该研究得到中国科学院B类先导科技专项、国家自然科学基金等项目的资助。
